Next-generation neural computations
Brains are not like computers. Our brains can quickly and easily spot familiar objects, like keys in a messy room, with very little effort. In contrast, even the best computers struggle to do this as fast or efficiently. This difference shows just how much more we need to learn about how our brains work to create smarter artificial intelligence.
To bridge the gap between neuroscience and Artificial Intelligence (AI), I seek to harness the efficiency of vision by understanding how neural computations govern sensory processes like vision and behavioral responses like eye movements.
Follow me on
![]() Mastodon.
Mastodon.
Biography
Laurent Perrinet is a computational neuroscientist (DR2 CNRS) at the Institut de Neurosciences de la Timone (UMR 7289, CNRS / Aix-Marseille Université), within the NeOpTo team. His research investigates predictive processing in the visual system — from single cortical cells to active vision and behavior — and its translation into neuromorphic algorithms. He has co-authored more than 63 peer-reviewed articles (h-index 30), supervised 6 completed PhD students and currently directs 3 PhD students (Alexandre Lainé, Matthis Dallain, Kevin Mairot). His work combines neurophysiology (Neuropixels recordings in marmoset), computational modeling (spiking neural networks, Free-Energy Principle) and open-source algorithmic development (MotionClouds, AnEMo, LogGabor).
- Predictive Processing & Active Inference
- Spiking Neural Networks & Neuromorphic Computing
- Computational Vision & Eye Movements
Habilitation à diriger des recherches, 2017
Aix-Marseille Université
PhD. in Cognitive Science, 2003
Université P. Sabatier, Toulouse, France
M.S. in Engineering, 1998
SupAéro, Toulouse, France
Lastest Publications
Recent Events

We are recruiting a PhD student to work on neuromodulatory control of predictive processing in mouse vision, co-supervised by Ede Rancz (INMED) and myself.
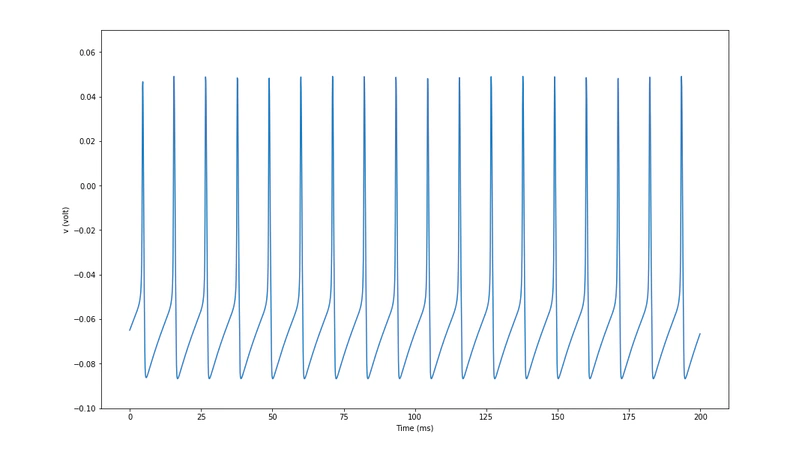
Project Overview
This CENTURI project combines:



Projects



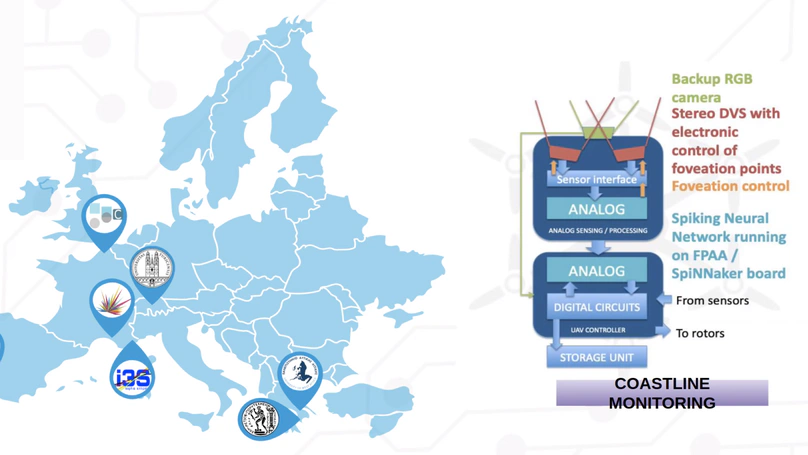
This would not be possible without…
Mentors
Current Students
Matthis Dallain
Phd candidate in Computational Neuroscience
Alexandre Lainé
Phd candidate in Computational Neuroscience
Agathe Choplin
PhD candidate in Neuroscience
Charlotte Roy
PhD candidate in Neuroscience
Kevin Mairot
Phd candidate in Ophtalmology
Collaborators
Ede Rancz
Researcher in Neuroscience
Andrew Isaac Meso
Lecturer, King’s College London (IOPPN).
Jonathan Vacher
Maître de Conférence (Associate Professor) @ MAP5, Université Paris-Cité.
Rick A Adams
Future Leaders Fellow, Institute of Cognitive Neuroscience & Centre for Medical Image Computing, UCL
Former Students
Antoine Grimaldi
PostDoc Researcher in Computational Neuroscience with Andrea Alamia in Toulouse, France
Thomas Kronland-Martinet
Post-doctoral Researcher
Jean-Nicolas Jérémie
Phd in Computational Neuroscience
Hugo Ladret
Postdoctoral Researcher at Friedrich Miescher Institute for Biomedical Research
Victor Boutin
CNRS researcher at CerCo (Toulouse, France).
Adrien Fois
Post-doctoral Researcher
Amélie Gruel
Post-doctoral researcher in neuromorphic computer science in Bordeaux, France
Angelo Franciosini
Biomedical Engineer @ Avicenna.AI.
Alberto Arturo Vergani
Post-doctoral Researcher @ Sant’Anna School of Advanced Studies, Pisa, Italy.
Wahiba Taouali
Consulting manager @ Enthought, Cambridge, United Kingdom.
Kiana Mansour-Pour
Executive DirectorExecutive Director, Shotise
Jean-Bernard Damasse
MD, Marseille, France.
Mina A Khoei
Senior AI/ML scientist @ SynSense, Zurich, Switzerland.
Jens Kremkow
PI @ Neuroscience Research Center, Charité, Berlin, Germany.
Nicole Voges
PostDoc in Computational Neuroscience
Hilde Langengen-Teigen
Phd candidate in Neuroscience
Jenna Fradin
Research Engineer at ISIR (Institut des Systèmes Intelligents et de Robotique)
Publications
Recent & Upcoming Talks
Grants





















Contact
How to reach me
- laurent.perrinet@univ-amu.fr
- +33 619 478 120
- Institut de Neurosciences de la Timone (UMR 7289), Aix Marseille Université - CNRS, Faculté de Médecine - Bâtiment Neurosciences, 27, Bd Jean Moulin, Marseille, PACA 13385 Marseille Cedex 05
- When you reach the INT building, take the stairs to the second floor, then enter the open space on the left and follow it to the end of the room; my office is on the right-hand side.
- OrcID
- mastodon
- Bluesky
- openAlex
- ResearcherID
- NeuroTree
- Google Scholar
- Zotero
- Publons
- arXiv
- GitHub
- pixelfed
- stackoverflow
- LastFM